
fV rámci výskumu sme sa zamerali na objektívne zhodnotenie prirodzeného zmladenia lesných drevín a stavu bylinného podrastu na rôzne narušených lesných biotopoch. Prirodzené zmladenie je základnou podmienkou lesných ekosystémov na ich pretrvávanie v čase. Jednoducho povedané, čo sa nedokáže obnoviť, skôr či neskôr zanikne. Bylinný podrast zase veľmi citlivo reaguje na zmeny podmienok prostredia (mikroklíma, pôda, konkurencia) a preto je využívaný na indikáciu týchto zmien. Vzhľadom na to, že cieľom bolo zhodnotiť schopnosť prirodzenej regenerácie lesných porastov po veternej kalamite v roku 2004, zaznamenávané bolo len zmladenie, ktoré nebolo staršie ako štyri roky. Celý výskum prebiehal v dolnej polovici Tichej doliny (obr. 1) na 16 rôznych výskumných plochách (obr. 2).
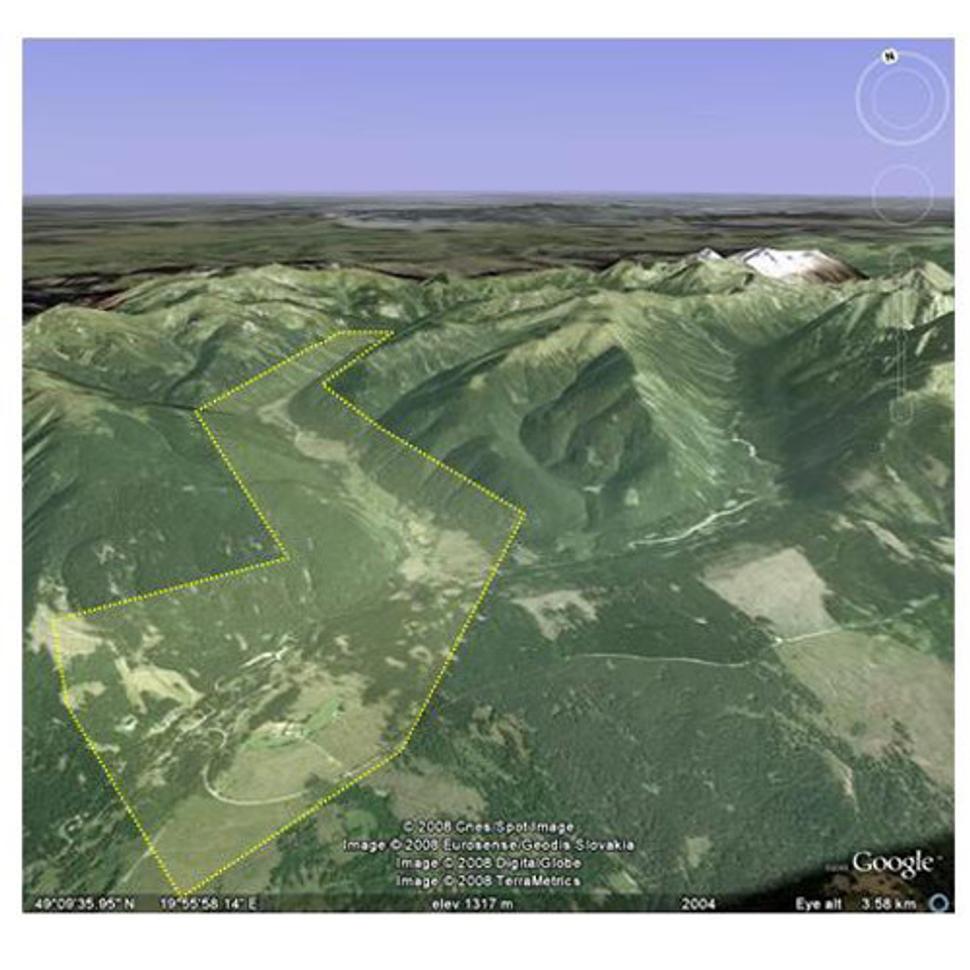
Obr. 1 Záujmové územie v Tichej doline
Sledovali sme štyri rôzne kategórie narušenia lesných porastov, z ktorých každá sa vyskytovala v štyroch nezávislých opakovaniach. Hodnotili sme stav sledovaných charakteristík v nenarušených lesných porastoch (S, obr. 3), v stojacej lykožrútovej kalamite (B, obr. 4) vzniknutej pôsobením silnej populácie lykožrútov po roku 2006, na nespracovaných polomoch (W, obr. 5), ktoré vznikli počas veternej kalamity v roku 2004 a na spracovaných kalamitných územiach (M, obr. 6), ktoré vznikli asanovaním následkov veternej a lykožrútovej kalamity po roku 2004.
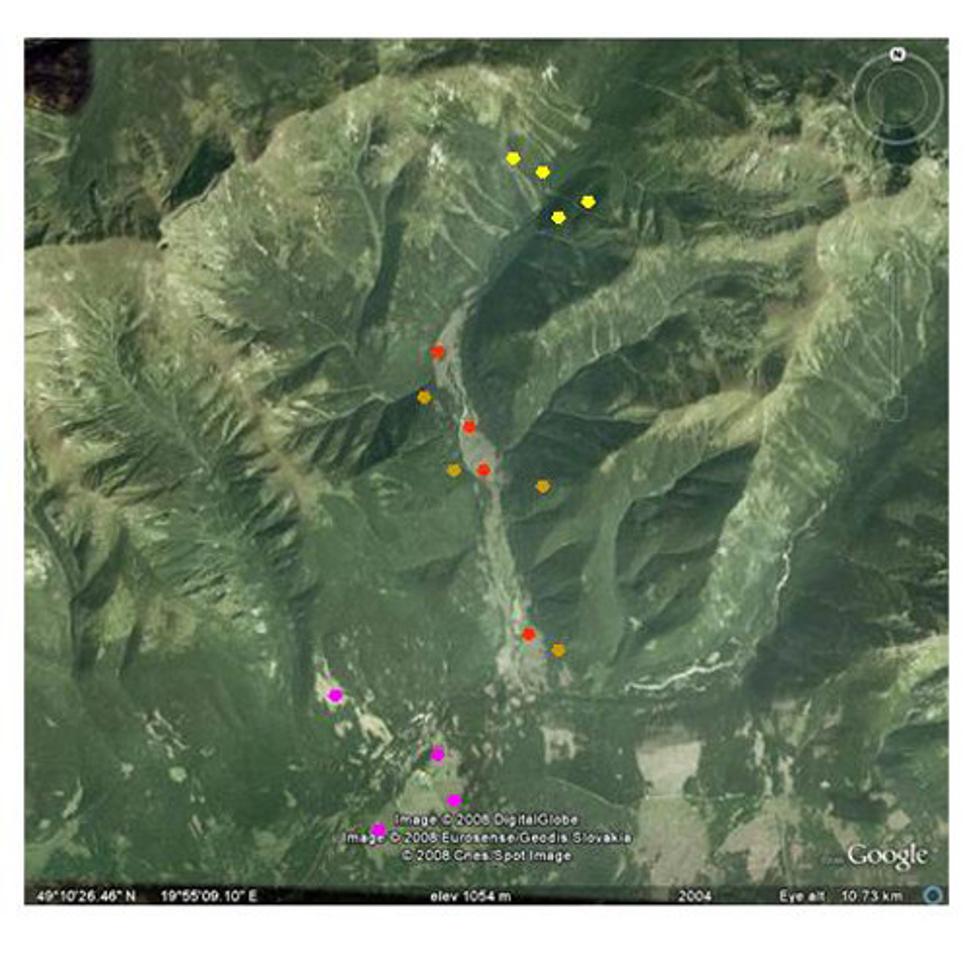
Obr. 2 Rozmiestnenie študijných plôch (žlté body - nenarušené smrekové porasty, oranžové - stojaca lykožrútová kalamita, červené - vetrové polomy, ružové - spracované kalamity).
V rámci každej sledovanej plochy sme v línii vytýčili 20 kvadrátov, každý s plochou 1m2 a rozostupom 3m, na ktorých sme spočítali prirodzené zmladenie drevín a odhadli pokryvnosť dominantne zastúpených bylín, ktorých celková pokryvnosť v rámci kvadrátu presahovala 25%. Celkovo sme tak zhodnotili zmladenie a stav bylinného podrastu na 320 kvadrátoch.

Obr. 3 Nenarušená horská smrečina s čučoriedkou (foto: T. Lepeška)

Obr. 4 Stojaca lykožrútová kalamita

Obr. 5 Nespracovaný polom

Obr. 6 Aktívne mamažované kalamitné územie (foto: T. Lepeška)
Takto získané údaje boli predmetom štatistického testovania, ktoré slúži na objektívne zhodnotenie trendov v dátových súboroch. Inými slovami, štatistické metódy nám umožňujú s konkrétne stanovenou určitosťou povedať, či je niečo také, alebo onaké. Bez emócií, subjektívnych pocitov a skresľovania reality.
Prirodzené zmladenie
V rámci výskumu sme zaznamenali významné zmladenie len v prípade jarabiny vtáčej (Sorbus aucuparia) a smreka obyčajného (Picea abies). Pri obidvoch drevinách sme zistili významné rozdiely medzi rôzne narušenými biotopmi (obr. 7).
Jarabina takmer úplne chýbala v nenarušených porastoch, avšak po narušení (vetrom alebo prostredníctvom lykožrúta) veľmi intenzívne zmladzovala, zjavne v dôsledku zmenených svetelných podmienok. Hustota populácie predstavovala priemerne takmer 40 000 jedincov na ha v prípade stojacej kalamity, a až 70 000 jedincov na ha na nespracovaných polomoch. Na spracovaných polomoch, bolo zmladenie jarabiny významne nižšie, len asi 10 000 jedincov na ha.
V okolí zmladenia neboli zaznamenané dospelé jarabiny, ktoré by mohli byť zdrojom semien. Jarabina patrí medzi dreviny, ktorých semená sú prenášané prostredníctvom vtákov a cicavcov (tzv. endozoochória). Vzhľadom na veľké množstvo mladých stromov, ktoré evidentne vyklíčili po narušení pôvodného porastu, je pravdepodobné, že semená jarabín prežívali v neaktívnom stave v pôde už pred kalamitou, priparavené vyklíčiť v prípade narušenia.
V prípade smreka bolo výrazné zmladenie zaznamenané v rámci všetkých nespracovaných biotopov, naopak na spracovaných polomoch prirodzené zmladenie tejto dreviny bolo takmer nulové. Denzita populácie na nespracovaných a nenarušených biotopoch sa pohybovala od 40 000 po 50 000 jedincov na ha. Aj keď v rámci hodnotenia celkového zmladenia smreka, neboli rozdiely medzi nenarušeným smrekovým porastom a kalamitnými plochami významné, pri hodnotení rôznych vekových kategórií smrekového zmladenia, sme zistili podstatné odlišnosti (obr. 8-10).
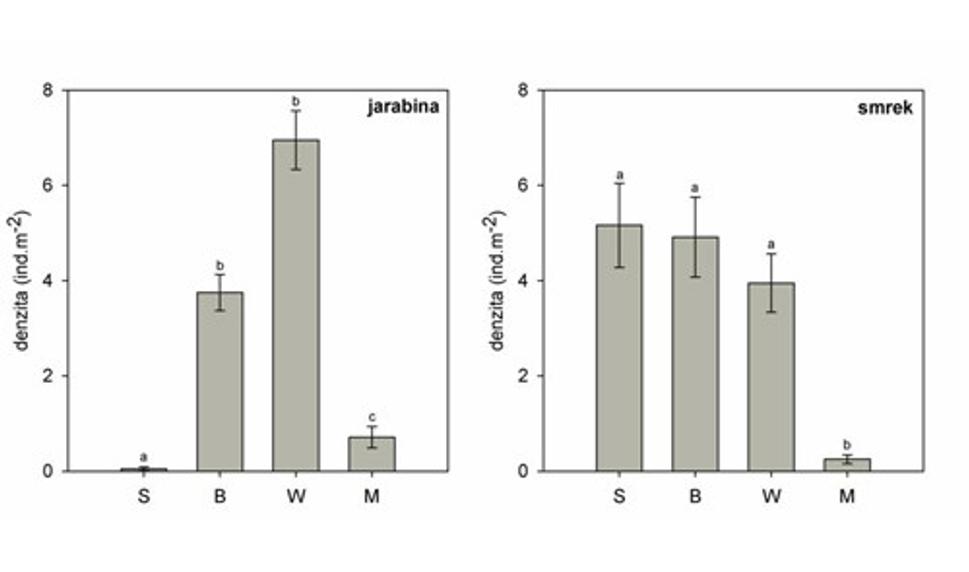
Obr. 7 Priemerná denzita zmladenia jarabiny vtáčej a smreka obyčajného v rámci štyroch postdisturbančných režimov. Rôzne písmena nad chybovými úsečkami označujú štatisticky významné rozdiely (p < 0,05) . (S-nenarušený smrekový porast, B - stojaca lykožrútová kalamita, W - nespracovaný polom, M - spracovaná kalamita).
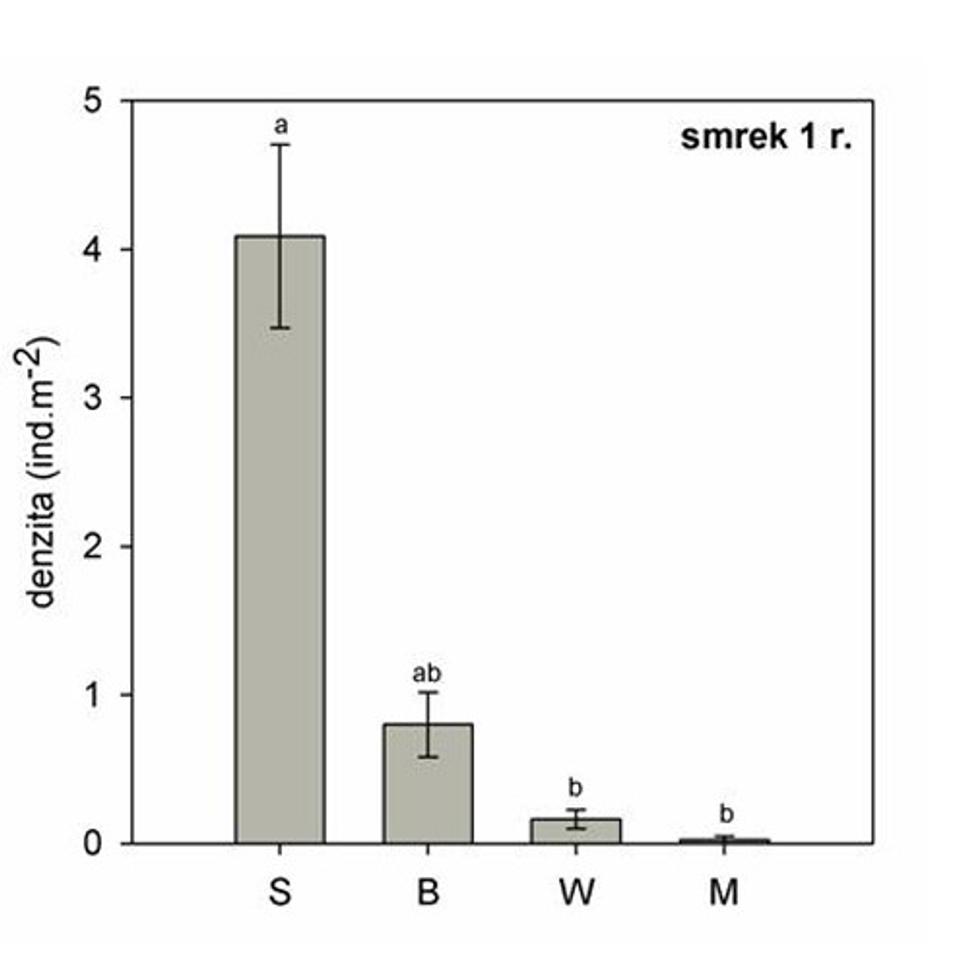
Obr. 8 Priemerná hustota jednoročných semenáčikov smreka v rámci štyroch postdisturbančných režimov. Rôzne písmena nad chybovými úsečkami označujú štatisticky významné rozdiely (p < 0,05) . (S-nenarušený smrekový porast, B - stojaca lykožrútová kalamita, W - nespracovaný polom, M - spracovaná kalamita).
V podmienkach nenarušeného smrekového porastu, patrí väčšina zmladenia do kategórie jednoročných smrekových semenáčikov (Obr. 8). Je to generácia smreka, ktorá vznikla v roku 2008, zo semien pochádzajúcich z dospelých stromov materského porastu. Tieto semenáčiky rastú v podmienkach kompaktného zápoja smrekového porastu. Z tohto dôvodu, nemajú potrebné množstvo svetla pre fotosyntézu a rast, a ich úmrtnosť je veľmi vysoká. V podstate po vyčerpaní živín zo semena, nie sú schopné aktívnej asimilácie látok z okolia a masovo hynú. V ďalšom vegetačnom období sú nahradené novou generáciou semenáčikov. Predstavujú svojím spôsobom pohotovostnú rezervu, ktorú každoročne vytvára materský porast, pre prípad narušenia.
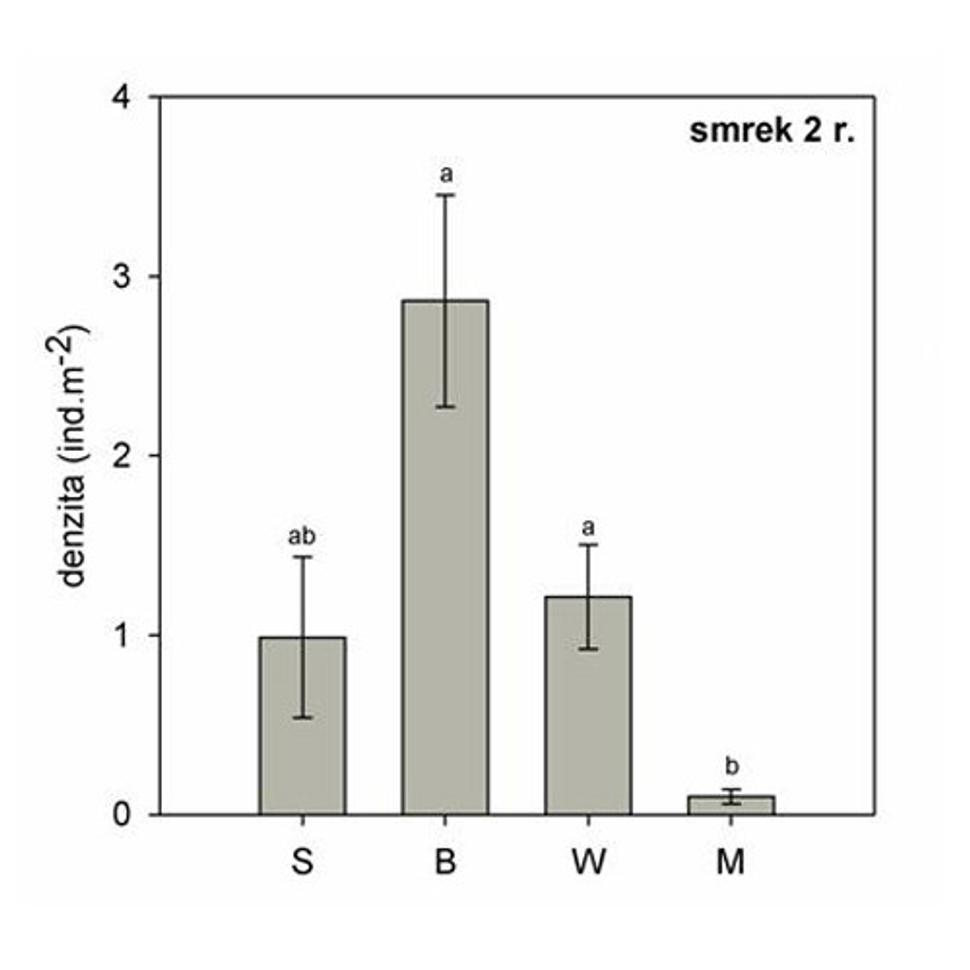
Obr. 9 Priemerná hustota dvojročného smreka v rámci štyroch postdisturbančných režimov. Rôzne písmena nad chybovými úsečkami označujú štatisticky významné rozdiely (p < 0,05) . (S-nenarušený smrekový porast, B - stojaca lykožrútová kalamita, W - nespracovaný polom, M - spracovaná kalamita).
Zmladenie v stojacej kalamite patrilo najmä do kategórie dvojročných smrekov (Obr. 9). Toto zmladenie pochádza zo semenáčikov z prelomu rokov 2006/2007. V tomto roku došlo k odumretiu materského porastu vplyvom premnoženia lykožrúta, čo malo za následok zmenu svetelných podmienok. Tieto umožnili prežitie semenáčikov smreka a ich rast. Odumretie materského porastu sa zároveň prejavilo na poklese denzity jednoročných semenáčikov, ktoré v porovnaní s predošlým biotopom dosahujú len asi štvrtinovú hodnotu.
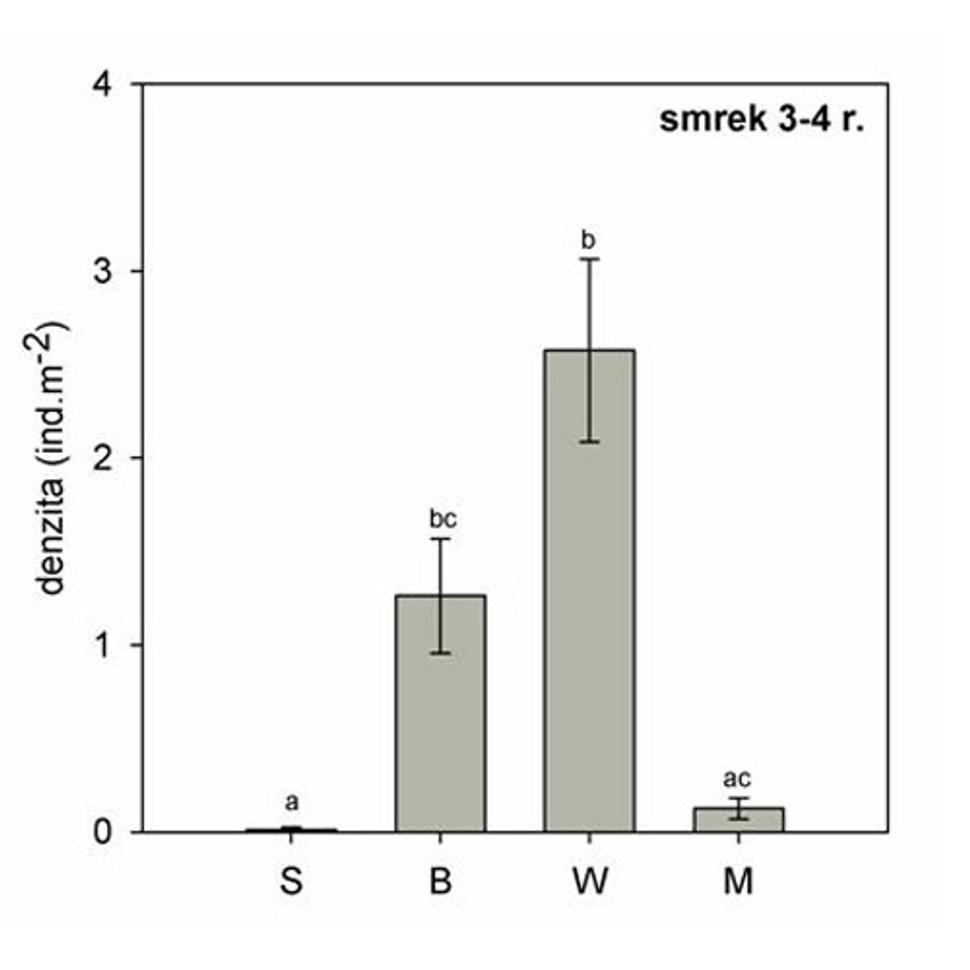
Obr. 10 Priemerná hustota troj- až štvorročných smrekov v rámci štyroch postdisturbančných režimov. Rôzne písmena nad chybovými úsečkami označujú štatisticky významné rozdiely (p < 0,05) . (S-nenarušený smrekový porast, B - stojaca lykožrútová kalamita, W - nespracovaný polom, M - spracovaná kalamita).
Najdlhšie narušenie materského porastu je typické pre nespracované polomy, na ktorých sú, v zmysle vyššie spomenutých zákonitostí, najviac zastúpené troj- až štvorročné smreky (obr. 10). Naopak jednoročné semenáčiky úplne chýbajú. Mladé smreky sa vyvíjajú v podmienakch polotieňa padnutých kmeňov, vo vzájomnej konkurencii s bylinným podrastom a zmladením jarabiny. Vytvárajú tak mozaikovitú štruktúru, tak potrebnú pre stabilitu dospelého porastu.
Bylinný podrast
Bylinný podrast nie je významný len z hľadiska indikácie zmien podmienok. Zohráva veľmi podstatnú úlohu v modikifikácii podmienok stanovišťa, a v súvisloti so zmladením drevín, predstavuje významný ekologický faktor, ktorý môže ovplyvniť schopnosť prežitia mladých stromčekov. Z tohto dôvodu sme v našom výskume hodnotili stav bylinných spoločenstiev v rôznych postdisturbančných režimoch.
Podobne ako v prípade prirodzeného zmladenia aj tu sme zaznamenali významné rozdiely v bylinnej vrstve (obr. 11). Bylinné spoločenstvá stojaceho lesa boli typické najmä výskytom machorastov. Boli významne odlišné od narušených nespracovaných spoločenstiev, a zároveň aj od spracovaných kalamít.
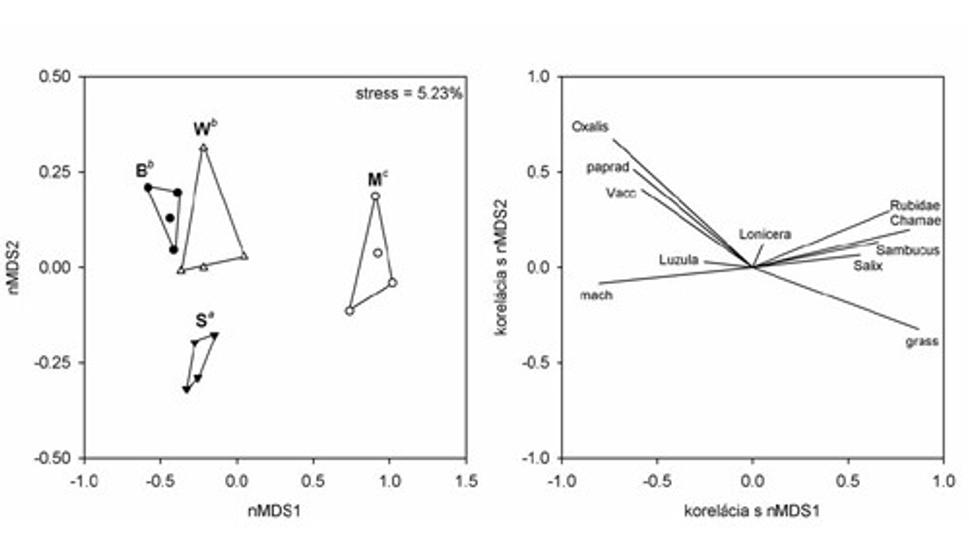
Obr. 11 Mnohorozmerná ordinácia postdisturbančných režimov na základe pokryvnosti dominánt prízemnej vegetácie. Rôzne písmena nad zhlukmi rôznych režimov označujú štatisticky významné rozdiely (p < 0,05) . (S-nenarušený smrekový porast, B - stojaca lykožrútová kalamita, W - nespracovaný polom, M - spracovaná kalamita).
Odklon od pôvodnej fytocenózy bol v prípade nespracovaných kalamitísk menší ako v prípade spracovaných. Aj keď rozdiel medzi režimami B a W bol v porovaní s nenarušenými porastmi významný, nešlo o kvalitatívnu zmenu. Poukazuje na to umiestnenie oboch nespracovaných režimov v ordinačnom priestore, kde sú oproti zhluku S rozdielne umiestnené len vo vzťahu k druhej osi. Takéto rozpoloženie je spôsobené najmä kvantitatívnymi zmenami v spoločenstve. Ľudskou rečou povedané, v narušených nespracovaných spoločenstvách rastie presne to isté, čo v nenarušenom lese, akurát je toho viac. Je to dôsledok odumretia stromovej vrstvy a zmeny svetelných podmienok.
Dôležité je však uvedomiť si, že aj napriek tomu, že stromy v narušených porastoch odumreli (stratili asimilačné orgány), ich drevná hmota je v ekosystéme stále prítomná a aktívne pôsobí. Z nášho hľadiska je momentálne najdôležitejšie to, že vytvára polotieň, ktorý umožňuje zachovanie prirodzenej skladby bylinných spoločenstiev.
Úplne opačná situácia nastáva v prípade spracovaných polomov. Tu nastal v porovnaní s nenarušeným porastom posun najmä na prvej osi ordinačného priestoru, s veľkosťou zhruba 1,5 jednotky, ktorá indikuje približne 75%-nú kvalitatívnu zmenu vo fytocenóze. Inými slovami, vo fytocenózach sa uplatňuje už len asi štvrtina pôvodných lesných druhov, ostatné boli nahradené inými druhmi (najmä trávy, vrbovka úzkolistá (Chamaerion angustifolium), a malina (Rubus idaeus). Zaujímavý je aj nástup pionierskych drevín ako baza červená (Sambucus racemosa), a vŕby (Salix spp.), ktoré boli zaznamenané výlučne v aktívne manažovaných spoločenstvách.
Najmä trávne druhy vystupujú ako veľmi významné modifikátory klímy holinových spoločenstiev, zároveň veľmi silne konkurujú pôvodným lesným druhom bylín ako aj smrekovému zmladeniu. Ich dominantné zastúpenie v rámci spoločenstva je umožnené aktívnym spracovaním kalamity, prostredníctvom ktorého je odstránená drevná hmota materského porastu a teda polotieň, typický pre nespracované plochy. Svoju úlohu určite zohráva aj mechanické narušenie pôdneho krytu s čím je spojené oslabenie pôvodných fytocenóz. Takto vzniknuté spoločenstvá tak predstavujú úplne iné štádium v rámci sukcesného radu - z regenerujúceho sa lesného spoločenstva (príklad nespracovaných polomov a stojacich kalamít) sa menia na pionierske prípravné spoločenstvo vysokých bylín a tráv.
Záver
Spracovanie akejkoľvek kalamity v rámci lesnej rezervácie je umelý zásah, ktorý vážne narúša lesný ekosystém a z prirodzene sa obnovujúceho lesa vytvára holinu závislú na umelom zalesnení. Nespracované kalamity sú naopak lesnými ekosystémami s vysokou regeneračnou schopnosťou, ktoré naďalej nesú podstatné znaky pôvodného lesného ekosystému. Naviac lesný porast, ktorý vzniká prirodzene po narušení, je zmesou drevín listnatých a ihličnatých - vďaka rôznorodosti podmienok nespracovanej kalamity je priestorovo a kvalitatívne diferencovaný a stabilný.
Tento záver je v zhode s naším výskumom, ale aj s predošlými výskumami z oblasti Šumavy a Bavorského lesa. Ponechanie kalamít na prirodzený vývoj preto v žiadnom prípade nepredstavuje nikým neoverený nebezpečný experiment, ani spôsob ako brániť lesu rásť. Je to spôsob, akým sa môžu horské smrekové biotopy vyvíjať spôsobom, na aký sú evolučne adaptované.






